Various cookies are used on our website: We use technically necessary cookies for the purpose of enabling functions such as login or a shopping cart. We use optional cookies for marketing and optimization purposes, in particular to place relevant and interesting ads for you on Meta's platforms (Facebook, Instagram). You can refuse optional cookies. More information on data collection and processing can be found in our privacy policy.
Einstieg in die neurologische Interdisziplinarität – Aufbau des Zentralnervensystems auf makroskopischer und zellulärer Ebene
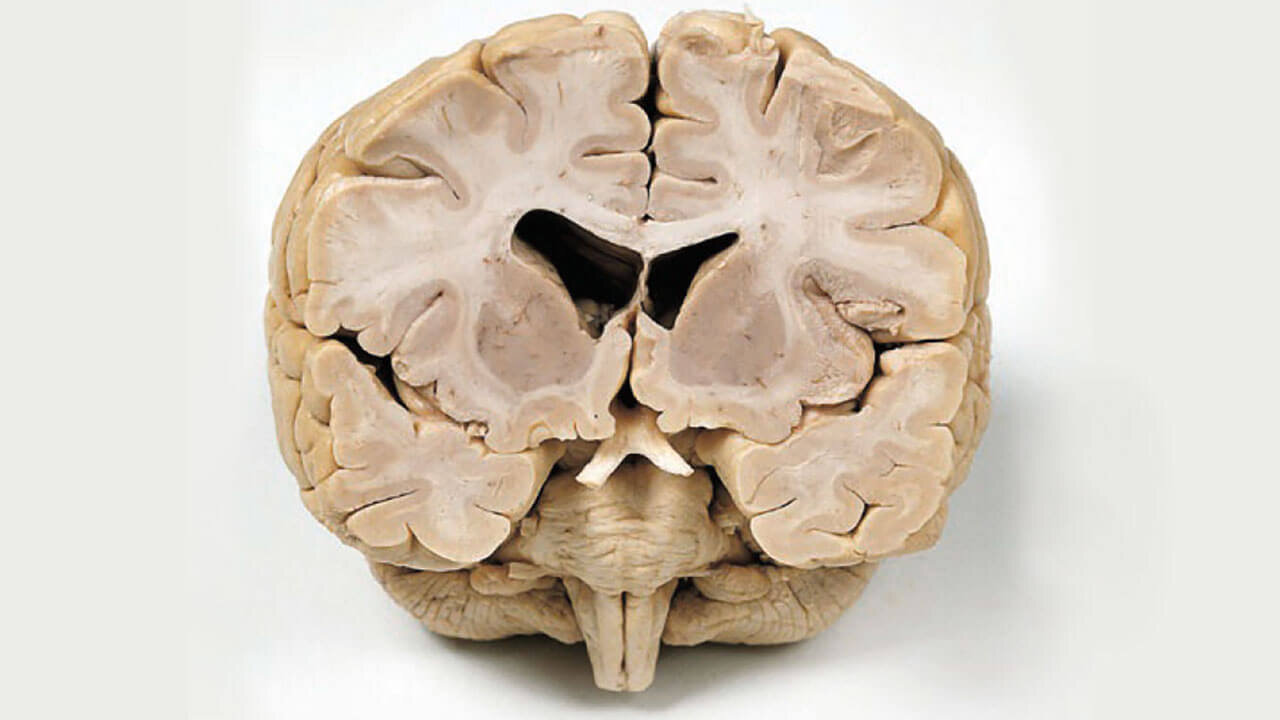
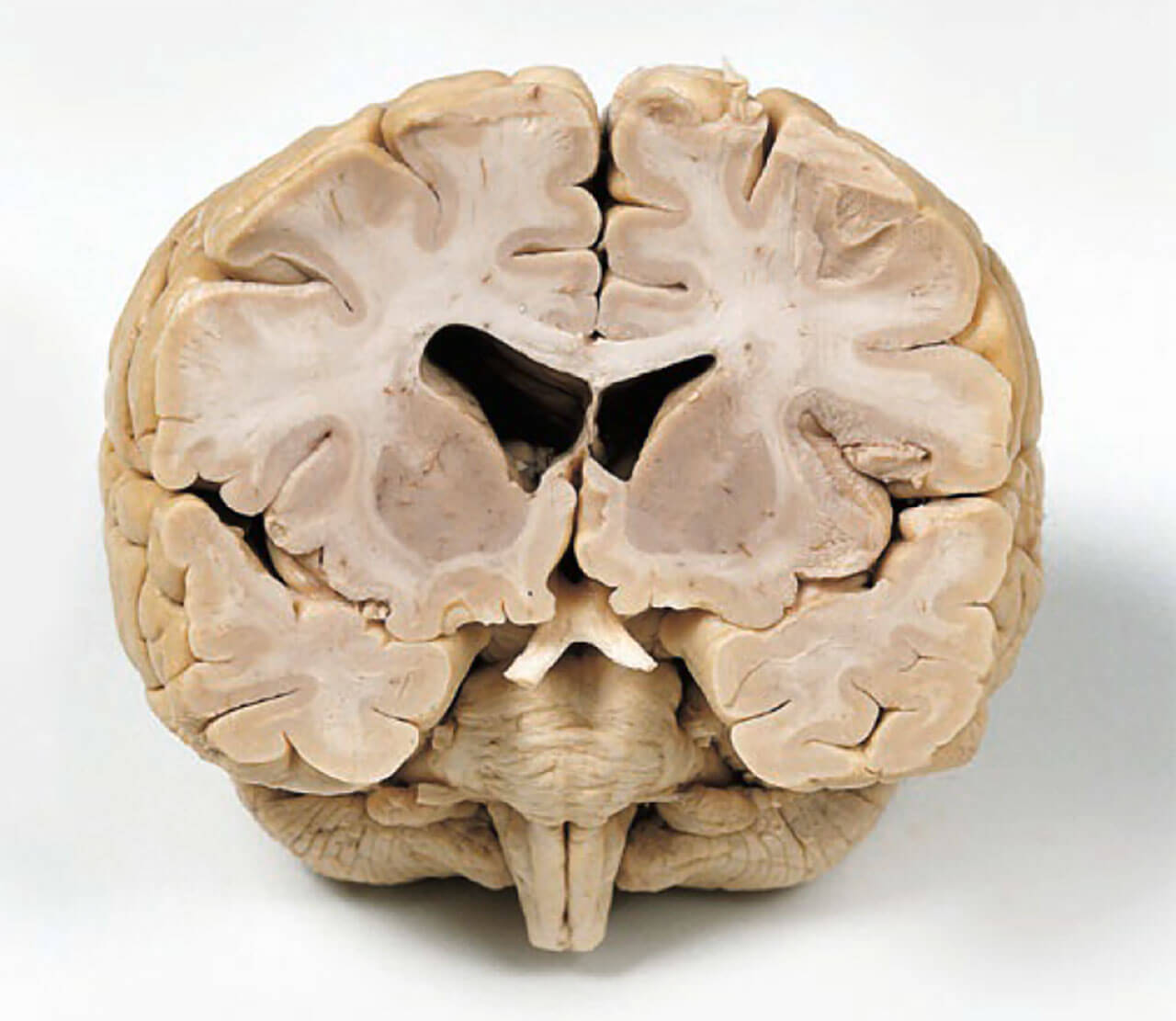
Frontalschnitt durch ein menschliches Gehirn. Die beiden mittig gelegenen „Löcher“ entsprechen den inneren Liquorräumen des Gehirns, den sogenannten Seitenventrikeln.
Das Nervensystem ist kompliziert und faszinierend zugleich. In keinem anderen wissenschaftlichen Feld konnten im vergangenen Jahrzehnt größere Fortschritte verzeichnet werden als in den Neurowissenschaften. Aus klinischer Sicht stellen uns neurologische und psychiatrische Erkrankungen mitunter vor große Herausforderungen, da deren Pathogenese nur ansatzweise verstanden wird und somit bisher keine kurative Behandlung zur Verfügung steht.
Im „Team Journal – Präventionsmedizin und Oralprophylaxe“ wird das Fachwissen vermittelt, das ZMP, DH, Zahnärztinnen und -ärzte und alle Fortbildungswilligen für einen erfolgreichen Arbeitsalltag brauchen. Besonders hervorzuheben ist der Fokus auf die Präventionsmedizin in der Rubrik „Interdisziplinär“, die den Patienten als Ganzes betrachtet. Mehr Infos zur Zeitschrift, zum Abo und zum Bestellen eines kostenlosen Probehefts finden Sie im Quintessenz-Shop.
Ein grundlegendes Verständnis des makroskopischen und mikroskopischen Aufbaus des Nervensystems, insbesondere von Gehirn und Rückenmark, ist entscheidend für die Entwicklung neuer therapeutischer Ansätze und für das Verständnis bereits zugelassener Therapieoptionen. Genau darum soll es in diesem Beitrag gehen: über den Aufbau des Zentralnervensystems auf makroskopischer und zellulärer Ebene.
Das Zentalnervensystem und das periphere Nervensystem
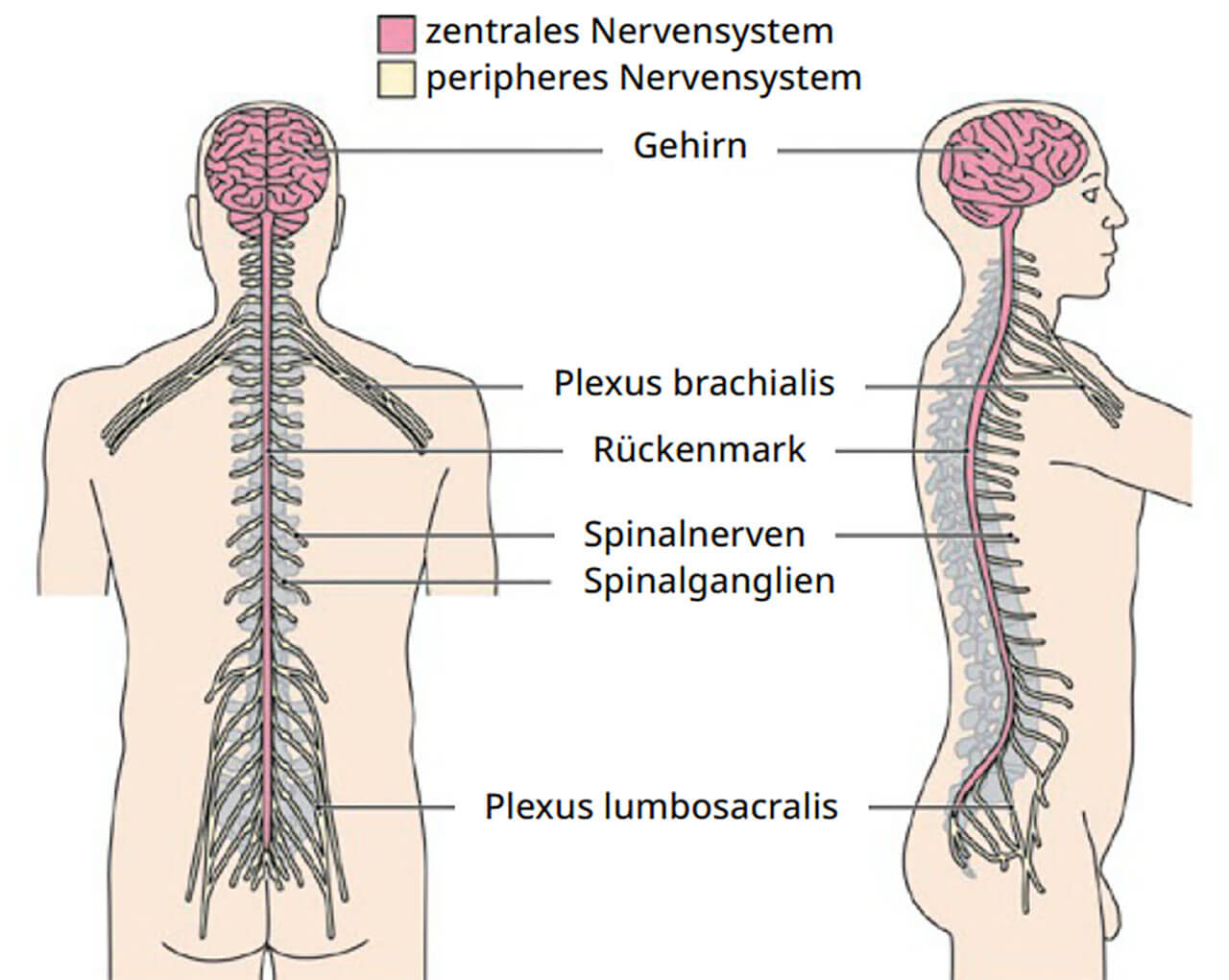
Auf den ersten Blick können, auf makroskopischer Ebene, zwei Anteile des Nervensystems unterschieden werden (Abb. 1): das Zentralnervensystem (kurz: ZNS) und das periphere Nervensystem (kurz: PNS). Das ZNS liegt, umgeben von knöchernen Strukturen, geschützt innerhalb des Schädels und der Wirbelsäule. Das ZNS ist nichts anderes als ein Hochleistungscomputer, der es uns ermöglicht, Reize und Signale aus unserer Umwelt zu empfangen, zu verarbeiten und adäquat auf die Reize zu reagieren. Hochentwickelte Abschnitte des ZNS, die an der Oberfläche des Gehirns, der sogenannten Hirnrinde zu finden sind, erlauben es uns Menschen, hochkognitive Aufgaben zu meistern – wie beispielsweise zu rechnen, zu reden oder aber Empathie und Abneigung zu zeigen. Das PNS liegt außerhalb des Schädels und des Wirbelkanals und hat vor allem die Aufgabe, sensible Impulse dem ZNS zuzuleiten beziehungsweise motorische Impulse des ZNS an die Muskulatur weiterzugeben. Komplizierte Rechenoperation sind dem PNS fremd.
Abb.1 Übersicht über die verschiedenen Anteile des Nervensystems. Das Gehirn und das Rückenmark sind Elemente des Zentralnervensystems. Spinalnerven, Spinalganglien, periphere Nervengeflechte und Hirnnerven (hier nicht gezeigt) werden dem peripheren Nervensystem zugerechnet.
Abb. 2 Frontalschnitt durch ein menschliches Gehirn. Die beiden mittig gelegenen „Löcher“ entsprechen den inneren Liquorräumen des Gehirns, den sogenannten Seitenventrikeln.
Als die alten Anatomen die ersten Gehirne inspizierten, begnügten sie sich natürlich nicht mit der Untersuchung des Gehirns von außen. Sie entschlossen sich dazu, das Gehirn zu sezieren um Informationen darüber zu erhalten, wie dieses faszinierende Organ von innen aussieht. Abbildung 2 zeigt ein menschliches Gehirn, welches von vorne angeschnitten worden ist. Neben den beiden zentralen Hohlräumen, die wir später noch als die inneren Liquorräume kennenlernen werden, fällt auf, dass manche Gebiete grau, andere weiß erscheinen. Die grau erscheinenden Bereiche nennt man graue Substanz (Substantia grisea), diejenigen, die heller erscheinen, nennt man weiße Substanz (Substantia alba). Der Streifen grauer Substanz, der das Gehirn umgibt, wird Hirnrinde (Kortex) genannt. Dort sitzen die neuronalen Zellkörper, die wichtig für hochkognitive Prozesse sind, zum Beispiel das Sprechen, Hören und Fühlen.
Zellulärer Aufbau des Nervensystems
Wir wollen uns nun ein wenig mehr dem zellulären Aufbau des Nervensystems widmen. Wir beschränken uns hierbei im Wesentlichen auf das ZNS. Prinzipiell können zwei Arten von „Hirnzellen“ unterschieden werden: die Nervenzellen, auch Neurone genannt, und die Gliazellen. Der Mitentdecker dieser zweitgenannten, nichtneuronalen Zellpopulation, Rudolf Virchow, vermutete Mitte des 19. Jahrhunderts eine Stütz- und Haltefunktion und gab ihnen deshalb den Namen Gliazellen, abgeleitet aus dem griechischen Wort „glia“ für Leim. Gliazelle ist also ein Sammelbegriff für strukturell und funktionell von den Neuronen abgrenzbare Zellen im Nervengewebe. Mithilfe unterschiedlicher Färbemethoden durch Santiago Ramón y Cajal, Pío del Río Hortega und Camillo Golgi konnten Gliazellen Ende des 19. Jahrhunderts weiter subklassifiziert werden. Beginnen möchten wir hier jedoch mit dem Aufbau einer Nervenzelle.
Aufbau und Funktion einer Nervenzelle
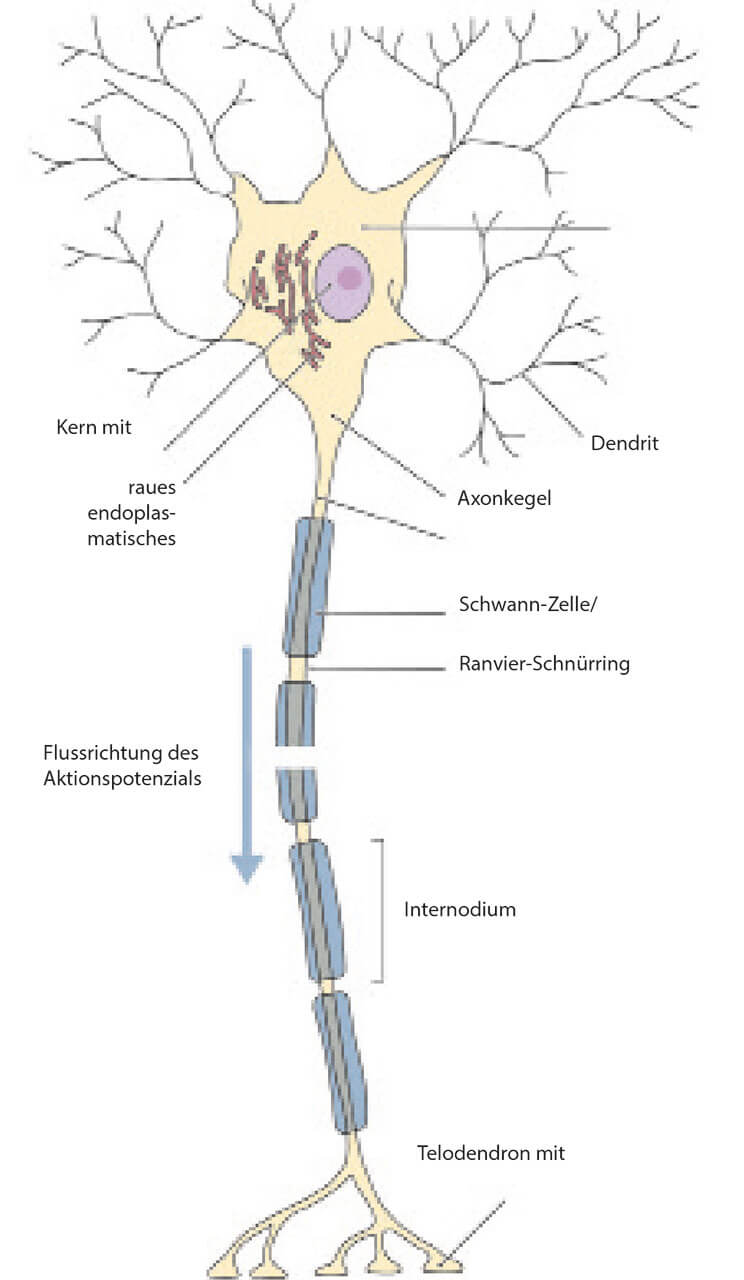
Die Grundlage jeglicher neuronaler Funktion ist, auf zellulärer Ebene, die Kommunikation zwischen Nervenzellen. Diese findet in Form von elektrischen Impulsen, sogenannten Aktionspotenzialen, statt. Neurone bestehen aus zwei unterscheidbaren Anteilen: einem Zellkörper (auch Soma oder Perikaryon genannt), der den Zellkern enthält, und zahlreichen dünnen Fortsätzen, die vom Zellkörper abgehen (Abb. 3). Diese Fortsätze bezeichnet man als Dendriten und Axone, die oft unter dem Oberbegriff „Neuriten“ zusammengefasst werden. Der elektrische Informationsfluss innerhalb einer Nervenzelle ist gerichtet: Dendriten nehmen Informationen mehrerer vorgeschalteten Nervenzellen an ihren Dendriten auf, verarbeiten die eingehenden Signale und leiten diese dann über ihre Axone in Form eines Aktionspotenziales zur folgenden Nervenzelle weiter. Der Informationsfluss, bezogen auf die Fortsätze der Nervenzelle, verläuft also von Dendrit über das Perikaryon zum Axon.
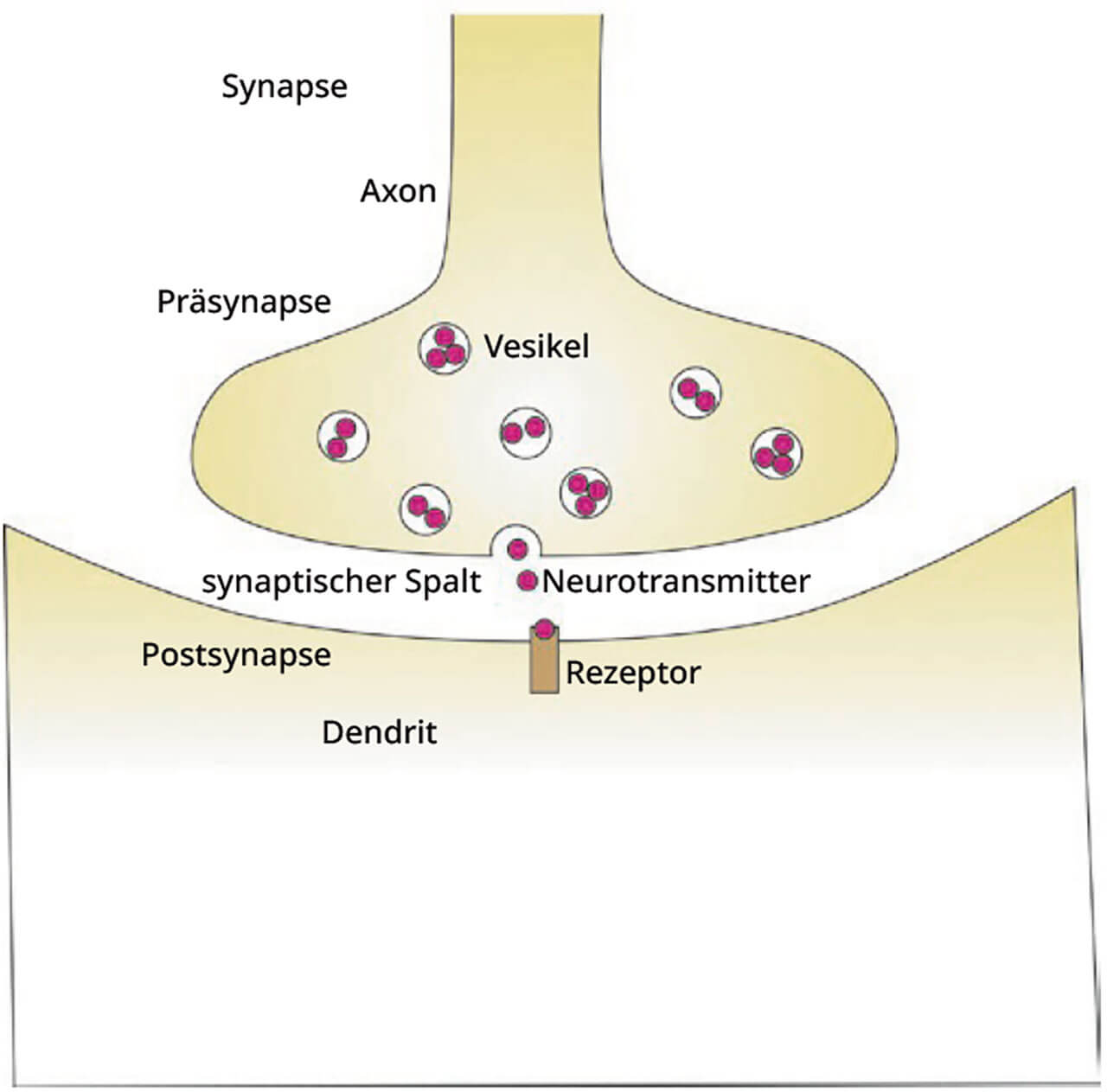
Der Ort der Kommunikation zweier Nervenzellen wird (chemische) Synapse genannt (Abb. 4). An deren Aufbau beteiligen sich das Ende eines Axons (Präsynpase) und eine Auftreibung (auch Dorn oder im Englischen „spine“ genannt) eines Dendriten der nachgeschaltenen Nervenzelle (Postsynpase).
Abb. 3 Übersicht über den prinzipiellen Aufbau einer Nervenzelle. Eine Nervenzelle besteht aus einem Nervenzellkörper (Soma/Perikaryon) mit zwei Arten von Fortsätzen (Neuriten): Dendriten, welche die Information aufnehmen, und Axone, welche die Information an die nächste Zelle weiterleiten. Ein ankommendes Aktionspotenzial wird an den Dornfortsätzen von einer Nervenzelle registriert. Am Axonhügel entsteht bei Überschreitung eines Schwellenwerts ein neues Aktionspotenzial. Dieses wird rasch über das (myelinisierte) Axon an die nächste Zelle weitergeleitet. Viele Axone sind von einer Myelinscheide umgeben; diese isoliert das Axon und beschleunigt somit die Fortleitung des Aktionspotenzials (saltatorische Erregungsleitung). An den Ranvier-Schnürringen ist die Myelinscheide regelmäßig unterbrochen. Dieser Bereich wird als Nodus bezeichnet, der Abschnitt zwischen zwei Ranvier-Schnürringen als Internodium. Zur besseren Orientierung ist die Flussrichtung der Information innerhalb einer Nervenzelle als Pfeil illustriert. An den Axonterminalen (synaptische Endköpfchen; Boutons) wird das Aktionspotenzial an die nächste Nervenzelle übergeben, man spricht von einer Verschaltung.
Abb. 4 Übersicht über den prinzipiellen Aufbau einer chemischen Synapse. Die bei der Weiterleitung eines Signals an die nächste Zelle beteiligten Strukturen fasst man als Synapse zusammen. Ein ankommendes Aktionspotenzial führt dazu, dass synaptische Vesikel, die Neurotransmitter beinhalten, mit der präsynaptischen Membran verschmelzen. Neurotransmitter werden dadurch in den synaptischen Spalt freigesetzt. Rezeptoren in der postsynaptischen Membran führen zur Erregung der postsynaptischen Zelle. Den gesamten Mechanismus der Weitergabe eines Aktionspotenzials nennt man synaptische Verschaltung.
Abb. 5 Übersicht über das prinzipielle Funktionsprinzip eines Netzwerks von Nervenzellen. Zum besseren Verständnis, wie die Kommunikation unter Nervenzellen funktioniert, und welche Rolle exzitatorische und inhibitorische Synapsen hierbei spielen, ein kleines bildhaftes Beispiel: Nehmen wir an, der Leiter einer Zahnarztpraxis muss entscheiden, ob ein neues Röntgengerät angeschafft wird oder nicht. Die Nervenzelle, welche diese Entscheidung treffen muss (wir nennen sie einmal Chef-Neuron), hat genau zwei Möglichkeiten. Entweder entscheidet sich das Chef-Neuron für oder gegen einen Kauf, verhält sich also binär, JA oder NEIN. Genauso funktioniert eine Nervenzelle auf Seite des Axons, genauer gesagt am Axonhügel, dem Ort, an dem das Axon vom Zellkörper entspringt. An dieser Stelle wird entschieden, ob ein oder kein Aktionspotenzial entsteht. Kommt ein Aktionspotenzial zustande, wird dieses über das Axon weitergeleitet. Nun werden Sie, als gute Chefin oder als guter Chef, eine solche Entscheidung womöglich nicht alleine fällen: Sie bitten ihre Mitarbeiter um deren Meinung. Einige, in der Abbildung in lila Farbe dargestellt, raten Ihnen vom Kauf eines neuen Röntgengeräts ab. Zu teuer, nicht wirtschaftlich, das Alte ist noch gut genug. Sie verhalten sich also, bezogen auf eine positive Kaufentscheidung Ihrerseits, inhibitorisch. Andere Kollegen wiederum raten Ihnen dringlich zum Kauf. Gerade gibt es tolle Fördermöglichkeiten, die Untersuchungszeiten verkürzen sich und neue Geräte können auch beginnende Gewebeveränderungen zuverlässig darstellen (in der Abbildung in grüner Farbe dargestellt). Diese Fürsprecher verhalten sich also, bezogen auf eine positive Kaufentscheidung Ihrerseits, exzitatorisch. Genau das passiert auf zellulärer Ebene in Nervenzellen: Afferente Nervenzellen wirken entweder erregend oder hemmend auf eine nachgeschaltete Nervenzelle, welche die eingehenden Informationen summiert, integriert und dann entscheidet, ob ein neues Aktionspotenzial gebildet wird oder nicht. Abbildung erstellt mit BioRender.com
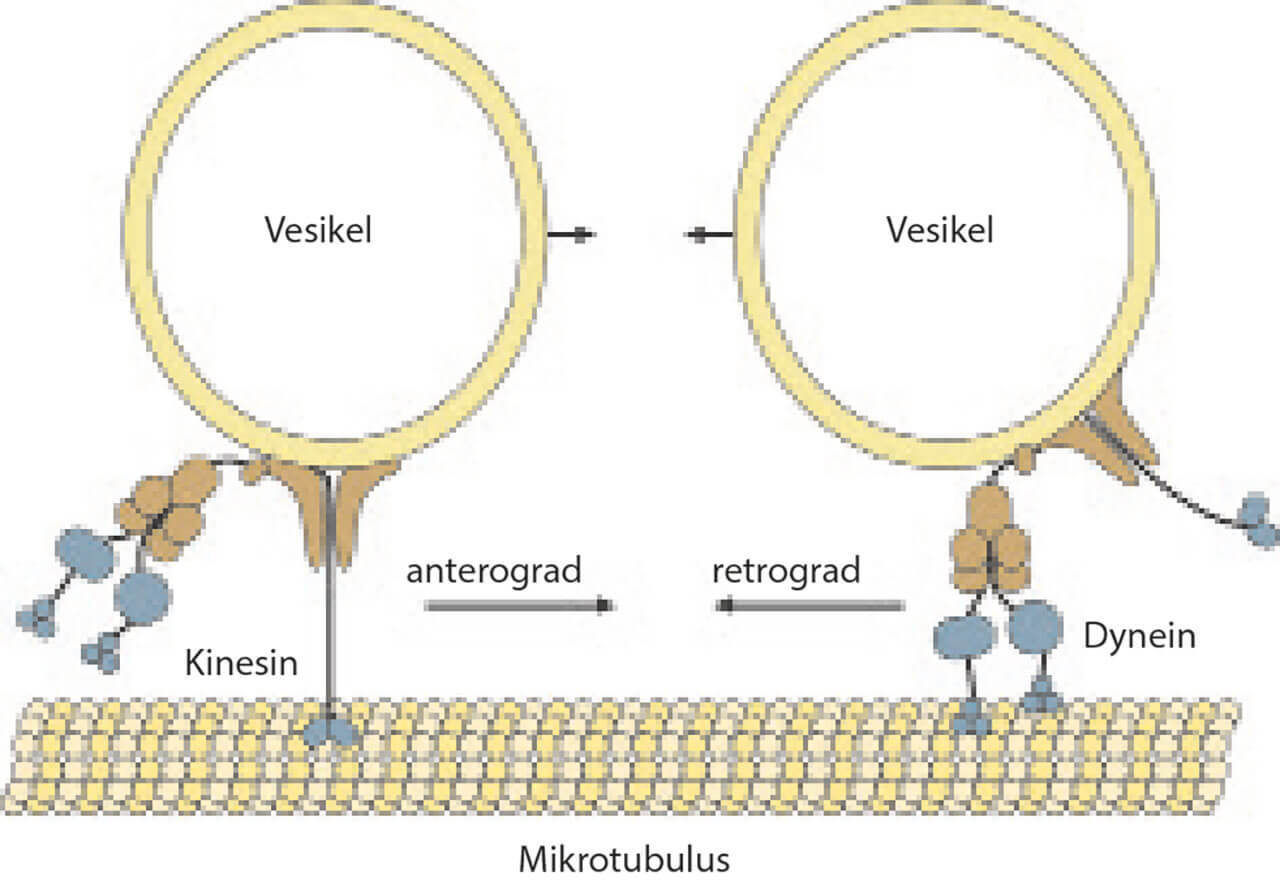
Abb. 6 Axone leiten nicht nur Aktionspotenziale weiter, sondern transportieren auch verschiedene intrazelluläre Substanzen (z. B. Neurotransmitter oder Wachstumsfaktoren). Man unterscheidet einen Transport in gleicher Richtung wie das Aktionspotenzial (anterograder Transport, vermittelt durch Kinesin) und einen in gegensätzlicher Richtung (retrograder Transport, vermittelt durch Dynein). Diese axonalen Transportvorgänge nutzen bestimmte Erreger aus, um sich im Gehirn einzunisten. Herpes-simplex- und Polioviren etwa gelangen durch den retrograden Transport ins Zentralnervensystem. Bei abgeschwächter Immunlage können schwerwiegende Entzündungen im Gehirn die Folge sein (Enzephalitis). Im Falle der Herpes-Viren können diese auch wieder entlang der Nervenbahnen in Richtung Haut wandern und dort zu einer lokalen Entzündung führen. Man bekommt schmerzhafte Herpesbläschen, z. B. im Bereich der Lippe.
Zwischen der Präsynpase und der Postsynpase befindet sich der synaptische Spalt. Die Übermittlung eines Aktionspotenzials zwischen zwei Nervenzellen heißt synaptische Übertragung beziehungsweise Verschaltung. Von den chemischen Synapsen des Nervensystems wird die elektrische Information im synaptischen Spalt in ein chemisches Signal umgewandelt, das den synaptischen Spalt überquert. An der postsynaptischen Membran wird dieses chemische Signal wieder in ein elektrisches umgewandelt. Das chemische Signal bezeichnet man als Neurotransmitter. Diese Neurotransmitter werden in synaptischen Vesikeln im Bereich der Präsynapse gespeichert und bei Bedarf von dort freigesetzt. Die über chemische Synapsen übertragenen Signale haben eine biochemisch festgelegte Wirkung.
Je nach Neurotransmitter und Ausstattung der postsynaptischen Membran, auf die das sendende Neuron Einfluss nimmt, wird entweder eine erregende (exzitatorische) oder aber eine hemmende (inhibitorische) Wirkung erzielt. Eine erregende Wirkung trägt dazu bei, dass die Zielzelle ein neues Aktionspotenzial am Axonhügel bildet, eine hemmende Wirkung wirkt gegensätzlich. Nicht nur einzelne Synapsen, sondern ganze Neurone werden daher in exzitatorische und inhibitorische unterteilt, je nachdem, ob sie erregende oder nur hemmende Synapsen an Zielzellen ausbilden. Zum besseren Verständnis, wie die Kommunikation unter Nervenzellen funktioniert und welche Rolle exzitatorische und inhibitorische Synapsen hierbei spielen, zeigt Abbildung 5 ein kleines, bildhaftes Beispiel.
Viele Substanzen werden im neuronalen Zellkörper (Soma/Perikaryon) synthetisiert und von dort über einen speziellen Transportmechanismus zu ihrem Zielort (zum Beispiel zur Synapse) transportiert: Man spricht vom axonalen Transport (Abb. 6). Vom Zellkörper zur Synapse (anterograd, stromabwärts) werden unter anderem Membranmaterial und zur Sekretion bestimmte Substanzen (wie Neurotransmitter) transportiert. Dies geschieht über Granula oder Vesikel, die an das Motorprotein Kinesin geheftet sind. Beim sogenannten retrograden Transport ist die Geschwindigkeit etwas geringer; hier werden Endprodukte des Stoffwechsels zurück zum Soma transportiert, außerdem zum Ab- und Umbau bestimmtes Membranmaterial sowie verschiedene Nervenwachstumsfaktoren, die für das Überleben der Nervenzelle notwendig sind. Der retrograde Transport erfolgt über Vesikel, die an das Motorprotein Dynein geheftet sind. Betrachten wir uns nun, in einem zweiten Schritt, die bereits erwähnten Gliazellen.
Gliazellen
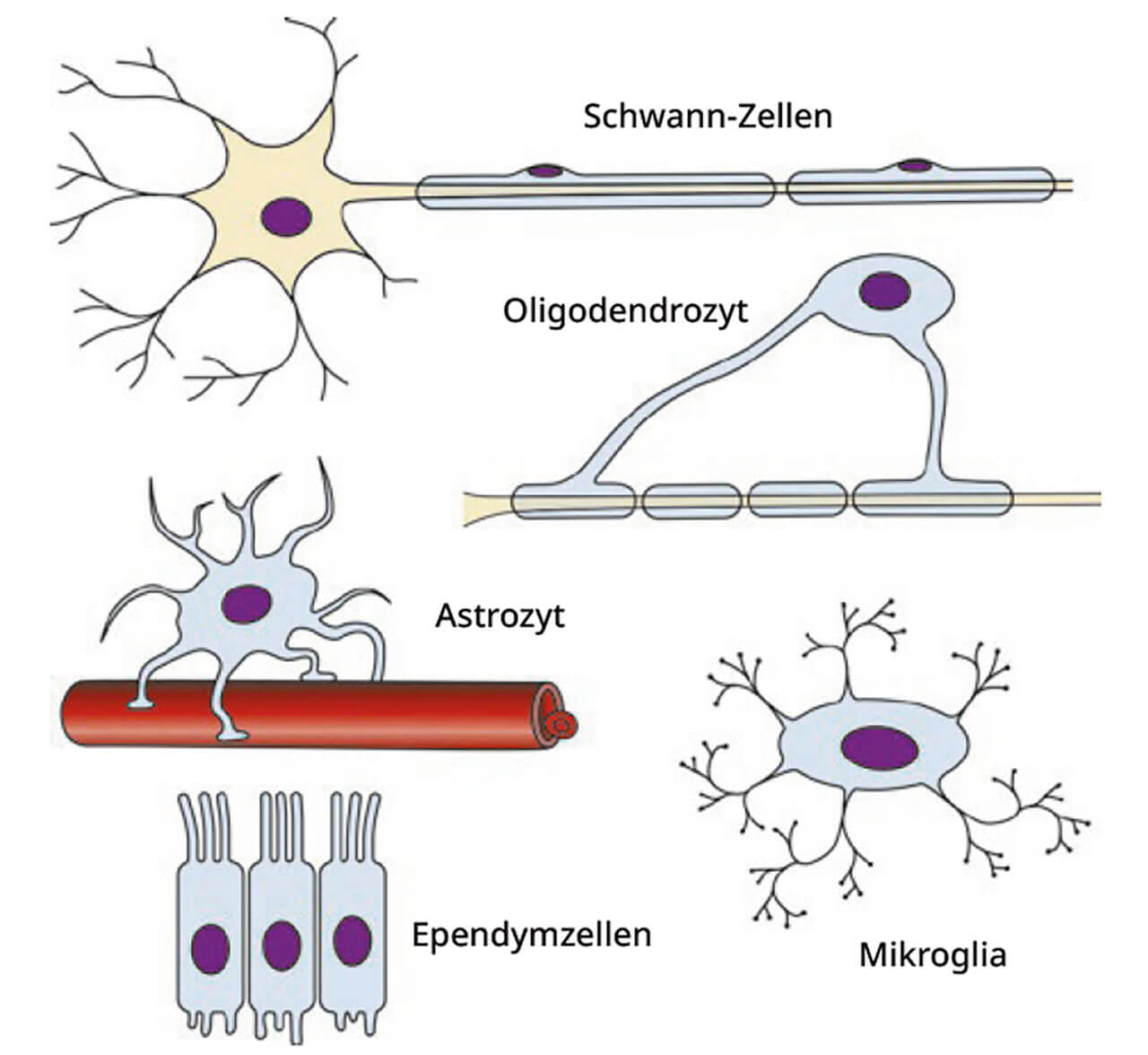
Abb. 7 Übersicht über verschiedene Gliazellen. Gliazellen lassen sich u. a. anhand ihrer Größe unterscheiden. Zu den Makrogliazellen des Zentralnervensytems zählt man Oligdendrozyten, die Myelinscheiden synthetisieren, Astrozyten, die unter anderem das chemische Milieu des Extrazellulärraums regulieren, und Ependymzellen, die die inneren Liquorräume auskleiden. Mikrogliazellen sind eine wichtige Effektorzellpopulation des angeborenen Immunsystems (Fresszellen = Phagozyten).Ersten Untersuchungen zufolge bilden Gliazellen ein Stützgerüst für die Nervenzellen (Astrozyten), sorgen für deren gegenseitige elektrische Isolation (Oligodendrozyten) und agieren als Fresszellen im Zuge von Infektionen und anderen neuronalen Pathologien (Mikroglia). Neuere Erkenntnisse zeigen, dass Gliazellen darüber hinaus maßgeblich am Stoff- und Flüssigkeitstransport sowie an der Aufrechterhaltung der Homöostase im Gehirn beteiligt sind und im Prozess der Informationsverarbeitung, -speicherung und -weiterleitung mitwirken1–5. Gliazellen sind somit unabdingbare Hilfszellen der Nervenzellen, deren spezifische Funktionen von ihnen abhängig sind. Sie besitzen Rezeptoren für Neurotransmitter und andere effektorische Moleküle. Es gibt sie sowohl im zentralen als auch im peripheren Nervensystem. Hier wollen wir uns auf die Zellen des ZNS beschränken.
Gliazellen können anhand ihrer Größe in Mikroglia von Makroglia unterschieden werden (Abb. 7). Markogliazellen werden weiter differenziert in Astrozyten (sternförmige Zellen), Oligodendrozyten (Zellen mit einigen wenigen Fortsätzen) und Ependymzellen.
Astrozyten
Bei Astrozyten handelt es sich um eine heterogene Population von Makrogliazellen mit einer Vielzahl von Funktionen. Empirische Daten aus verschiedenen Disziplinen der letzten zwei Jahrzehnte widerlegen die lang gehegte Annahme, dass die Informationsverarbeitung im Gehirn ausschließlich von Neuronen durchgeführt wird. Protoplasmatische Astrozyten, die dominierenden Zellen in der grauen Substanz von Säugetieren, sind für die normale synaptische Funktion und deren Erhaltung unverzichtbar. Zudem spielen sie eine entscheidende Rolle bei der Expression, Speicherung und Konsolidierung synaptischer Informationen, von einzelnen Synapsen bis hin zu globalen neuronalen Netzwerken6.
Um die Bedeutung der Astrozyten für eine regelhafte neuronale Funktion nachvollziehen zu können, lohnt sich ein Blick in die Evolutionsgeschichte. Das Volumen menschlicher protoplasmatischer Astrozyten, der häufigsten Zellen des Kortex, ist 27-mal größer und ihr Durchmesser 2,55-mal größer als bei Nagetieren. Darüber hinaus sind Astrozyten durch ein Netzwerk reichlich verzweigter peripherer Fortsätze gekennzeichnet. Sie umhüllen nahezu alle exzitatorischen Synapsen im Gehirn. Bemerkenswert ist, dass die Anzahl der Fortsätze menschlicher Astrozyten um den Faktor 10 höher ist als bei Nagetieren. Der menschliche Neokortex beherbergt außerdem mehrere anatomisch definierte Unterklassen von Astrozyten, die bei Nagetieren nicht vorhanden sind7.
Schnellere Kommunikation zwischen Astrozyten durch die Ausbreitung von Kalziumwellen
Interessanterweise ergaben mikroskopische Analysen, dass die Geschwindigkeit der Kommunikation zwischen Astrozyten durch die Ausbreitung von Kalziumwellen beim Menschen schneller ist als bei Nagetieren. Eine Anpassung der Astrozyten im Laufe der Zeit scheint demnach ein wichtiger Faktor für die evolutionäre Entwicklung der Spezies Mensch gewesen zu sein. Aus Platzgründen können nicht alle Funktionen von Astrozyten genannt werden, auf einige wichtige soll an dieser Stelle jedoch eingegangen werden.
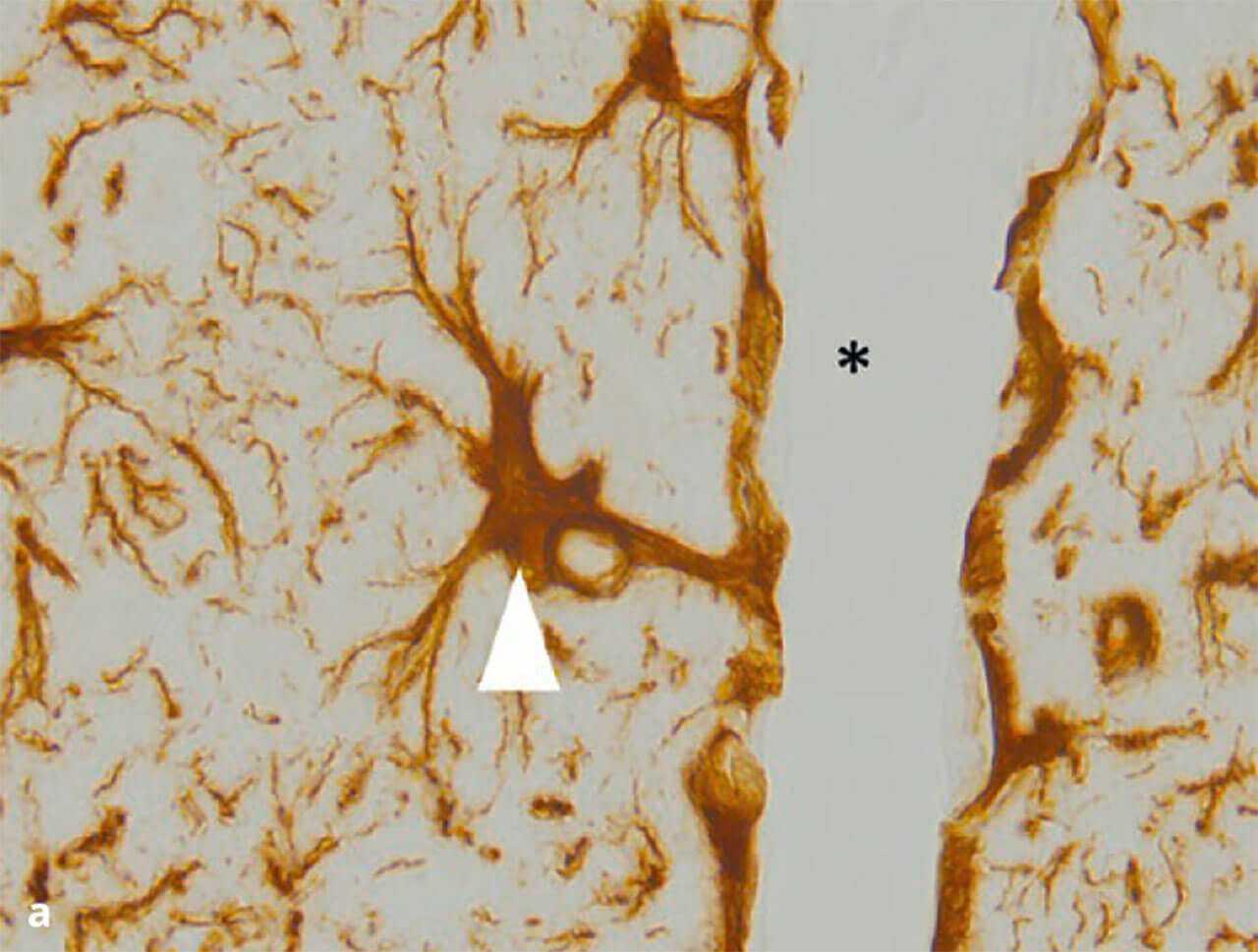
Astrozyten sind über Nexus („Gap junctions“) verbunden und besitzen viele verzweigte Fortsätze, von denen einige an Blutgefäßen enden und hier eine Schicht aus Gliafüßchen aufbauen, die sogenannte Membrana limitans gliae perivascularis (Abb. 8a). Sie ist am Aufbau und der Funktion der Blut-Hirn-Schranke beteiligt, einer Struktur, welche verhindert, dass Blutbestandteile ohne weiteres in das Gehirn eindringen können. Astrozyten der grauen Substanz sind darüber hinaus am Aufbau der sogenannten Membrana limitans gliae superficialis beteiligt (Abb. 8b). Diese Grenzmembran, aufgebaut aus einer dichten Schicht von astrozytären Zellfortsätzen und einer direkt daran angrenzenden Basalmembran, stellt die äußere Grenzfläche des Hirngewebes dar. Die Funktion der Membrana limitans gliae superficialis besteht darin, das Eindringen von Erregern von außen zu verhindern, was beispielsweise im Rahmen einer Hirnhautentzündung (Meninigitis) von großer Bedeutung sein kann.
Abb. 8a und b Astrozyten übernehmen viele Aufgaben im Gehirn. Man unterscheidet die der weißen (fibrillären) und der grauen (protoplasmatischen) Substanz. Unter anderem bilden sie isolierende Barrieren nach außen hin, entweder in Richtung Hirnoberfläche (a: Glia limitans superficialis) oder um Gefäße herum (b: Glia limitans perivascularis). Färbung gegen „Glial fibrillary acidic protein“ (GFAP). Pfeil in (a) = Zellkörper eines Astrozyten mit Ausbildung einer Glia limitans perivascularis. Stern in (a) = Lumen eines Gefäßes. Pfeil in (b) Glia limitans superficialis. Stern in (b) Oberfläche des Gehirns.
Abb. 8a und b Astrozyten übernehmen viele Aufgaben im Gehirn. Man unterscheidet die der weißen (fibrillären) und der grauen (protoplasmatischen) Substanz. Unter anderem bilden sie isolierende Barrieren nach außen hin, entweder in Richtung Hirnoberfläche (a: Glia limitans superficialis) oder um Gefäße herum (b: Glia limitans perivascularis). Färbung gegen „Glial fibrillary acidic protein“ (GFAP). Pfeil in (a) = Zellkörper eines Astrozyten mit Ausbildung einer Glia limitans perivascularis. Stern in (a) = Lumen eines Gefäßes. Pfeil in (b) Glia limitans superficialis. Stern in (b) Oberfläche des Gehirns.
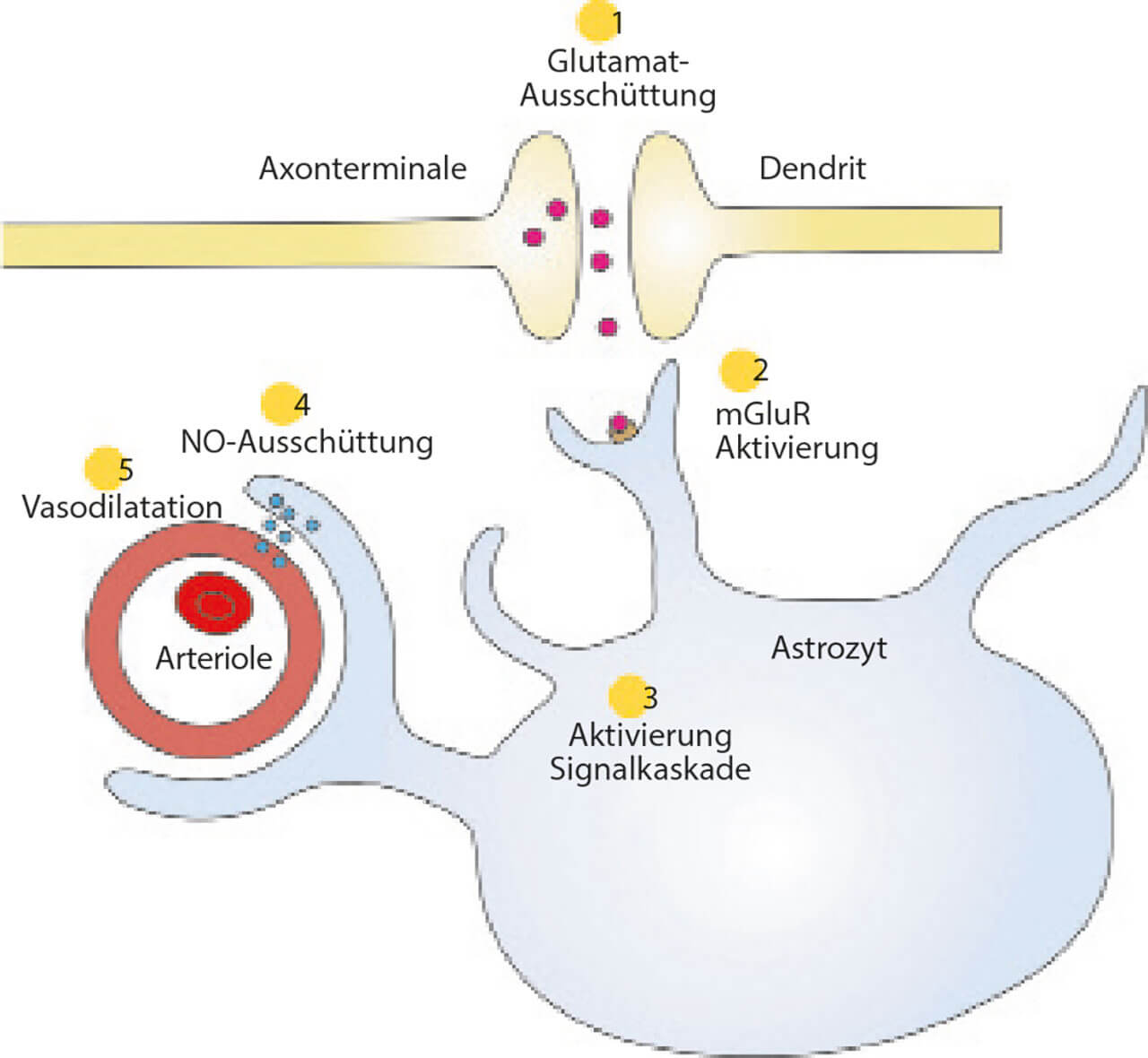
Abb. 9 Mechanismus der neurovaskulären Kopplung, vermittelt durch Astrozyten – Schritt 1: Glutamat wird freigesetzt; Schritt 2 Glutamat bindet u. a. an metabotrope Glutamat-Rezeptoren (mGluR) von umliegenden Astrozyten; Schritt 3: Dies bewirkt die Aktivierung einer Signalkaskade im Innern der Astrozyten; Schritt 4: Die Signalkaskade hat eine Ausschüttung von Stickstoffmonoxid (NO) an einem periarteriellen Fortsatz zur Folge; Schritt 5: NO ruft dort eine lokale Vasodilatation und somit eine Steigerung der Durchblutung hervor.
Darüber hinaus sind Astrozyten an der sogenannten neurovaskulären Kopplung beteiligt. Die neurovaskuläre Kopplung ist ein physiologischer Mechanismus zur Regulierung der Blutversorgung des Gehirns, um den Mehrbedarf von aktivem Nervengewebe an Sauerstoff und Glukose durch lokale Steigerung des Blutflusses zu decken (Abb. 9).
Astrozyten rücken immer mehr in das Interesse der Forschung, die Fortschritte sind mehr als vorzeigbar. Mit der Neuromyelitis-optica (auch optikospinale Enzephalomyelitis oder Devic-Syndrom) wurde jüngst ein Erkrankung identifiziert, der eine autoimmune Reaktion gegen ein astrozytäres Protein, dem Wasserkanal Aquaporin-4, zugrunde liegt. Typisch ist eine meist beidseitige Optikusneuritis (Entzündung des Sehnervs) in Kombination mit einer Myelitis (Entzündung des Rückenmarks) mit Querschnittssyndrom. Während das kranielle MRT in der Regel unauffällig ist, zeigt das spinale MRT meist einen langstreckigen Befall, der sich über mehr als drei Rückenmarkssegmente erstrecken kann.
Oligodendrozyten
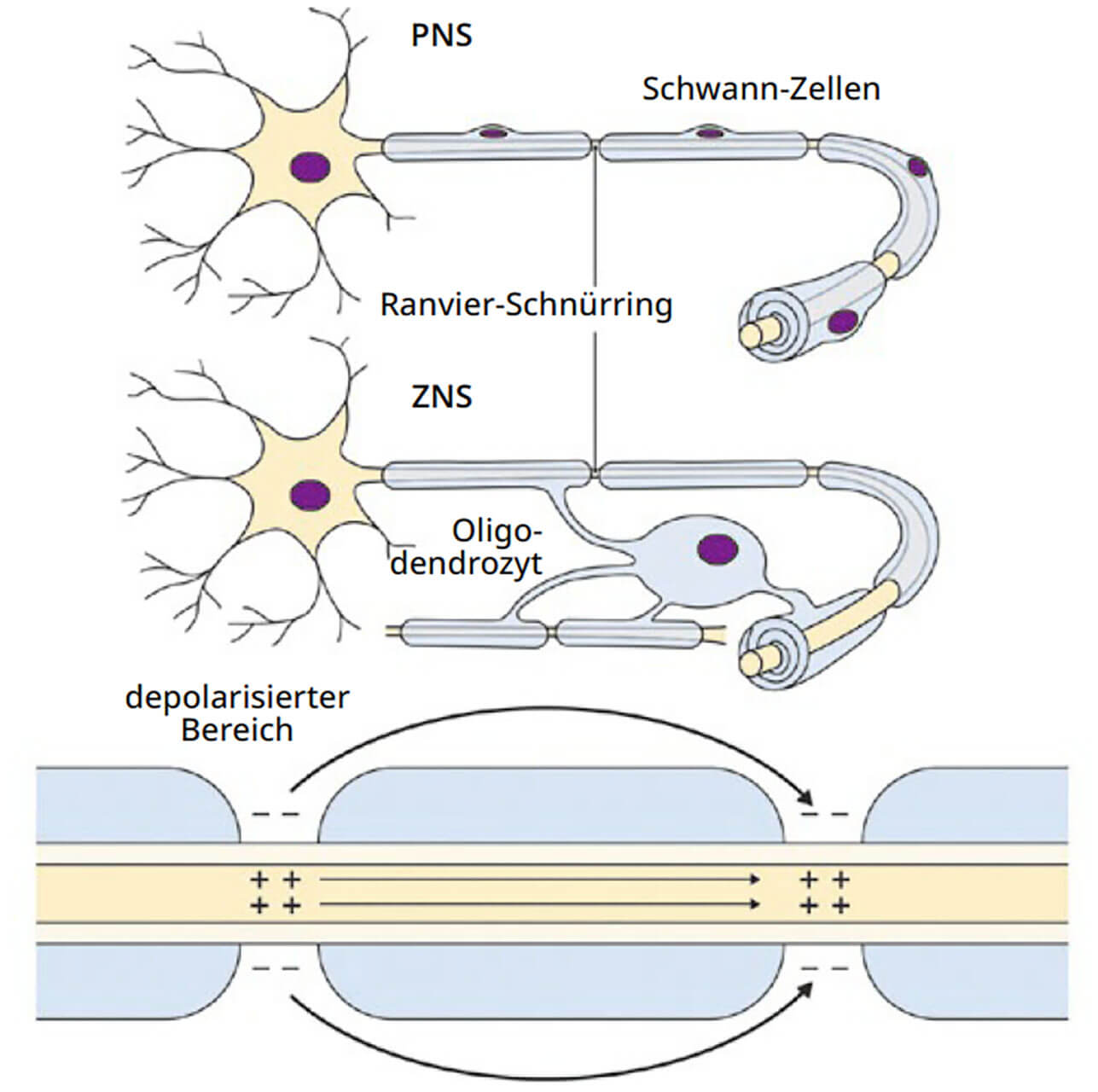
Oligodendrozyten und Schwann-Zellen bilden die Myelinscheide, eine lipidreiche Biomembran, welche die Axone der meisten Nervenzellen von Wirbeltieren spiralförmig umgibt und somit elektrisch isoliert. Oligodendrozyten sind die Myelinbildner innerhalb des ZNS, Schwann-Zellen bilden die Myelinscheiden der peripheren Nerven.Hier wollen wir uns, wie bereits im vorangestellten Abschnitt, mit dem ZNS beschäftigen, also den Oligodendrozyten.
Myelin wurde 1854 von dem Pathologen Rudolf Virchow (1821–1902) mithile der Lichtmikroskopie an Gewebeschnitten entdeckt. Er fand in Nervenfasern eine Markscheide und schlug vor, sie Myelin (griech. myelòs – „Mark“) zu nennen. Im Vergleich zu anderen Biomembranen weist Myelin einen besonders hohen Lipidgehalt (70 Prozent) und einen relativ geringen Proteinanteil (30 Prozent) auf. Daher erscheint Myelin in der makroskopischen Sicht weiß, weshalb stark myelinisierte Regionen im ZNS auch als „weiße Substanz“ bezeichnet werden, im Gegensatz zur gering myelinisierten „grauen Substanz“.
Myelinscheiden haben zwei zentrale Aufgaben. Zum einen isolieren sie die auf engstem Raum verlaufenden Axone und verhindern so, dass ein Aktionspotenzial von einem auf das andere Axon überspringt (verhindert also „Kurzschlüsse“). Zum anderen vergrößert eine Myelinscheide die Geschwindigkeit, mit der ein Aktionspotenzial entlang eines Axons weitergeleitet werden kann. Eine Steigerung der Leitungsgeschwindigkeit ist vor allem mit Hinblick einer schellen Verarbeitung neuronaler Impulse extrem wichtig. Um die Leitungsgeschwindigkeit zu erhöhen, hat die Evolution zwei unabhängige Mechanismen entwickelt. Ein Mechanismus besteht darin, den Axondurchmesser zu vergrößern, wie es zum Beispiel beim Tintenfisch der Fall ist. (Hier gilt: Je größer der Durchmesser eines Axons, desto schneller die Leitungsgeschwindigkeit.) Er beträgt hier fast einen Millimeter! Der zweite Mechanismus ist die Myelinisierung, also das Umwickeln des Axons mit den Membranen von Oligodendrozyten (Abb. 10).
Bei der Multiplen Sklerose, kurz MS, bilden sich autoreaktive Lymphozyten, die vermutlich gegen einen Bestandteil der Myelinscheide gerichtet sind. Charakterisiert ist diese Erkrankung durch wiederkehrende entzündliche Herde innerhalb des Gehirns, die von einem Untergang von Oligodendrozyten und einer Zerstörung der Myelinscheiden (Demyelinisierung) gekennzeichnet ist.
Abb. 10 In der Peripherie werden die Myelinscheiden von Schwann-Zellen produziert. Im Zentralnervensystem übernehmen dies die Oligodendrozyten. Die Myelinscheiden sind hierbei nicht kontinuierlich um ein Axon gewickelt, sondern entlang der Axone regelmäßig von den Ranvier-Schnürringen unterbrochen. Nur an den Ranvier-Schnürringen entstehen Aktionspotenziale, nicht aber in den myelinisierten Bereichen des Axons (Internodien). Dieser Aufbau ermöglicht die saltatorische Erregungsleitung, welche deutlich schneller als die kontinuierliche Erregungsleitung nichtmyelinisierter Fasern ist. Außerdem spart diese Art der Erregungsleitung Energie, da ein Aktionspotenzial nur am Ort der Schnürringe und nicht kontinuierlich entlang eines Axons aufgebaut werden muss.
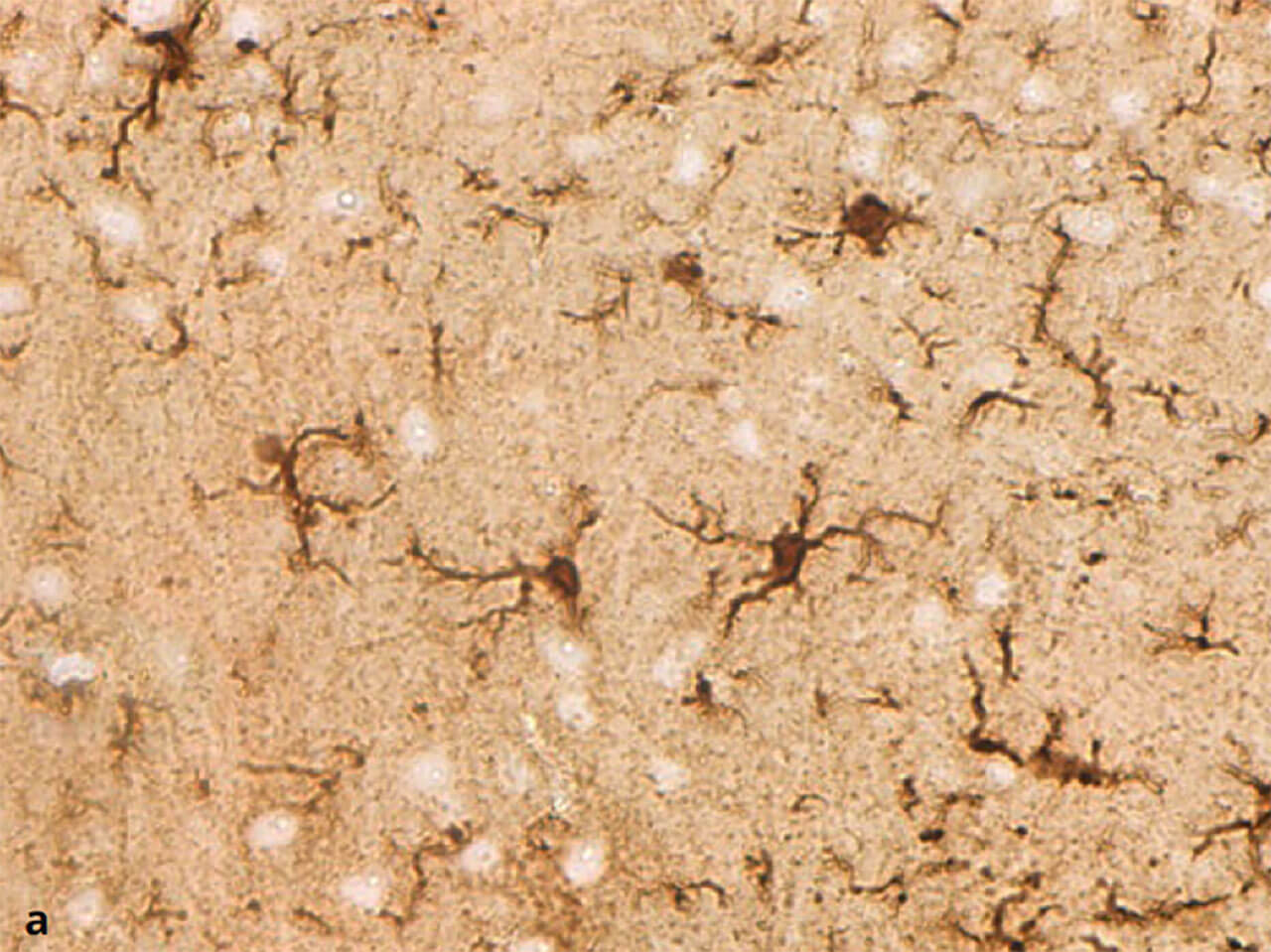
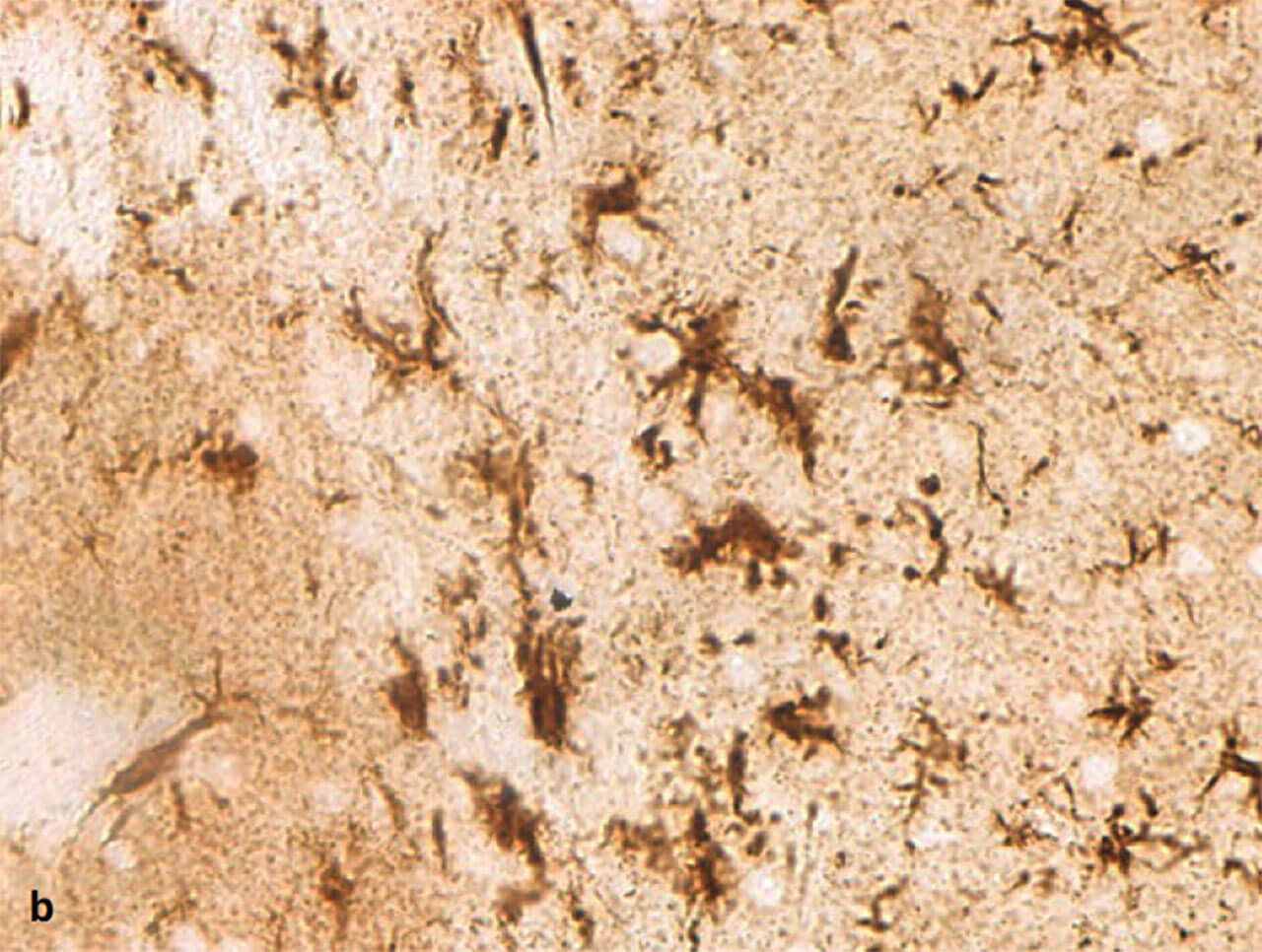
Abb. 11a und b Ruhende (a) und aktivierte (b) Mikroglia. Mikrogliazellen sind die Fresszellen des Zentralnervensystems. Ruhend haben sie einen kleinen Zellkörper und schlanke, verzweigte Fortsätze. Im aktivierten Zustand schwellen sie an und ziehen ihre Fortsätze ein (Färbung gegen das Protein IBA-1).
Abb. 11a und b Ruhende (a) und aktivierte (b) Mikroglia. Mikrogliazellen sind die Fresszellen des Zentralnervensystems. Ruhend haben sie einen kleinen Zellkörper und schlanke, verzweigte Fortsätze. Im aktivierten Zustand schwellen sie an und ziehen ihre Fortsätze ein (Färbung gegen das Protein IBA-1).
Mikroglia
Als Mikroglia oder Mesoglia bezeichnet man eine Gruppe von Immuneffektorzellen des ZNS. Sie werden zwar formal zur Familie der Gliazellen gerechnet, im eigentlichen Sinn handelt es sich jedoch um Zellen des mononukleär-phagozytären Systems. Ihre wichtigste Aufgabe besteht in der Phagozytose von Erregern sowie in der Ausschüttung entzündungssteigender Moleküle.Es wird davon ausgegangen, dass sie im Laufe der Entwicklung in das ZNS einwandern. Wie aus ihrem Namen bereits abgeleitet werden kann, handelt es sich bei der Mikroglia um die kleinste Gliazellpopulation. Mikroskopisch sieht man schmale, lang gestreckte Zellen, die einen irregulären, länglichen Zellkern mit dichtem Chromatin besitzen. Die Zellfortsätze können fein und sehr verzweigt sein (ramifizierte, ruhende Mikroglia; Abb. 11).
Ein Beitrag von Prof. Dr. Dr. Markus Kipp und Lukas Wenzel, beide Rostock
Our systems reports that you are using an active AdBlocker software, which blocks all page content to be loaded.
Fair is fair: Our industry partners provide a major input to the development of this news site with their advertisements. You will find a clear number of these ads at the homepage and on the single article pages.
Please put www.quintessence-publishing.com on your „adblocker whitelist“ or deactivate your ad blocker software. Thanks.
Hilfestellungen für die Praxen zur GOZ, Abgrenzung zur GOÄneu, Unterstützung junger Kolleginnen und Kollegen und Abgeber – Dr. Romy Ermler, Vizepräsidentin der BZÄK, zu Gast in Folge #29 von Dental Minds